
細胞膜を構成するリン脂質は二重膜の内と外で非対称になっているという話の続き。
特にフォスファチジルセリン(PS)については、表に出てくると殺されるというとてもロマンチックな機構が関わっていました。
これには酵素が絡んでいて、ホスファチジルセリン–フリッパーゼなんて名前がつけられていますが…大きな分子の移動って、もはや酵素の限界を超えていないか?というのが疑問でした。
ここで、Wikipediaよりフォスファチジルセリンの構造を見てみます。
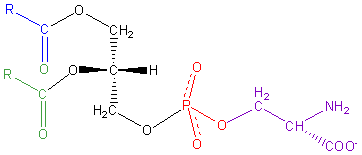
左のR-COO-は脂肪酸なので炭素鎖が連なる脂溶性部分で、前回の記事で用いた図でいうと黄色の二本足部分に相当します。
そして赤の水溶性部分に相当するのがリン酸+セリン。セリンは側鎖のOHで結合しています。
このような分子は生体内の水溶液中では、極性のため界面に集まったりミセルを作ったり、あるいは細胞膜のような構造をとらないと物凄く不安定です。
なので、この二重膜構造はリン脂質的には安定な構造であり、表と裏でのリン脂質の移動はそれなりのエネルギーが必要になるはず。
そこで酵素の登場となるわけですが…
引っかかるのは、「内側だけ」という局在と、これだけでかい分子をどうやって移動させるかの2点。
何でもできることで名を馳せている酵素ですが、さすがに能力の限界超えていないかと(^ー^;
だって化学反応を触媒するというのが酵素の役割のはずで、物質の移動とかってその範疇外では?
(ただし高校生物の範囲であっても、物質の移動を酵素が行っている事例はありました。ナトリウムポンプとか、グルコーストランスポーターとか。でも具体的な機構はそんなに明らかでもなかったような…)
しかし、現実には細胞はリン脂質をアシンメトリーに保っていることがわかっているようです。
「フリッパーゼ」でググると、そんなに古くない論文がいくつか引っかかるし、日本語でも記事がいくつか出てきました。
このフリッパーゼ、英語のウィキには触媒する反応がこんな簡単に書かれています。
ATP + H2O + phospholipid in ⇔ ADP + phosphate + phospholipid out
Phospholipid-translocating ATPase|Wikipedia
inとかoutってなんだよって話ですがw
EC番号を探ると、一応は加水分解酵素である3が振られていますが、トランスロカーゼという分類が新たに登場しているようで、分類作業もまだいじられてる最中の様子(^-^;和名はまだない。
細胞膜のアシンメトリーについては最新の研究テーマにもなっているような題材のようですので、どこまで明らかにされてるのかはまだ良くわかりません。
もう少し探ってみます。

